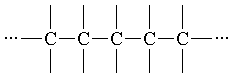
В процессе Развития Материи по оргуровню З постепенно формировались органические соединения, представлявшие собой все более динамически устойчивые фн. системы, которые, в свою очередь, затем становились фщ. единицами в системах более высшего порядка. К таким динамически устойчивым органическим соединениям можно отнести, в частности, аминокислоты. Общая формула их построения такова:
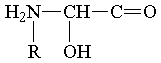
где R - фн. ячейка углеводородного радикала, которую могут занимать и другие различные фщ. единицы.
Из сотен и тысяч молекул аминокислот (как фщ. единиц)
синтезируются более сложные молекулы белковых веществ, или белков (фн. систем),
которые по истечении периода их функционирования под влиянием минеральных кислот,
щелочей или ферментов распадаются на составляющие их фщ. единицы - аминокислоты
с тем, чтобы дать им затем возможность вновь войти в состав образующихся новых
соединений, то есть заполнить соответствующие им новые фн. ячейки. И процесс этот
повторяется постоянно бесконечное число раз.
|
|
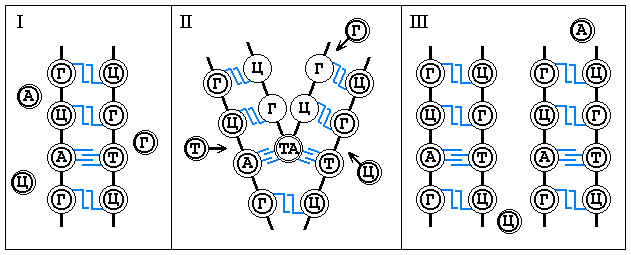
Из 104 элементов периодической системы Менделеева в клетках обнаружено более 60. Атомы кислорода, углерода, водорода и азота заполняют 98% фн. ячеек клеточных подсистем. 1,9% предоставлены атомам калия, серы, фосфора, хлора, магния, натрия, кальция и железа. Менее 0,1% фн. ячеек занято прочими веществами (микроэлементами). Различные сочетания указанных элементов дают несколько типов внутриклеточных подсистемных образований, которые каждая клетка включает в свои фн. ячейки в качестве фщ. единиц в следующих пропорциях (в %):
|
|
|||||||||||||||||||
Все указанные вещества, сами сложные в структурном
отношении, не нагромождены в клетке вместе в хаотическом беспорядке, а в качестве
фщ. единиц заполняют расположенные в строго определенном порядке предназначенные
для каждого из них фн. ячейки ее единой структуры. Функционируя, они проделывают
свои четко определенные микродвижения внутри микрообъема пространства клетки,
регулируемые соответствующими внутриклеточными алгоритмами, при этом существует
безусловная связь этих движений в пространстве как с абсолютным, так и с относительным
течением времени. Каждое из веществ клетки в качестве фщ. единицы несет строго
определенную функцональную нагрузку и имеет свои, регламентируемые соответствующими
алгоритмами, периоды функционирования. Все их разнообразное сочетание представляет
собой единый тонко отрегулированный клеточный механизм.
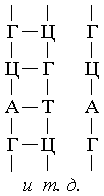
Графически это выглядит следующим образом:
Молекулы РНК также являются полимерами, как и ДНК,
но в отличие от них имеют одну спираль фн. ячеек, а не две. РНК выполняет в клетке
ряд функций, в том числе
 Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.
Переписывание информации на и-РНК, то есть процесс "транскрипции", происходит
при одновременном синтезировании и-РНК, которое осуществляется с помощью принципа
комплементарности. В результате образующаяся цепочка и-РНК по составу и последовательности
своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов
одной из цепей ДНК. Молекулы и-РНК направляются затем к рибосомам, куда поступают также
и аминокислоты, доставляемые извне клетки в уже готовом виде. Аминокислоты попадают
на рибосому в сопровождении транспортных РНК (т-РНК), состоящих в среднем из 70 - 80
нуклеотидных звеньев, в 4 - 7 местах комплементарных друг другу. К одному из концов
т-РНК присоединяется аминокислота, а в верхушке изгиба находится триплет нуклеотидов,
который соответствует по коду данной аминокислоте. На каждую аминокислоту имеется своя
т-РНК, то есть их также 20 разновидностей.![]()
[ Оглавление ] [ Продолжение текста ]